| HOME | HELP | FEEDBACK | SUBSCRIPTIONS | ARCHIVE | SEARCH | TABLE OF CONTENTS |
| ||||||||||||||||||||
Institution: WESTERN OREGON UNIV | Sign In via User Name/Password
|
|
|||||||||||||||||||||||||||||
Institution: WESTERN OREGON UNIV | Sign In via User Name/Password |
||||||||||||||||||||||||||||||
Population Biology |
2 Department of Plant Science, Unit 4067, University of Connecticut, Storrs, Connecticut 06269 USA 3 Department of Ecology and Evolutionary Biology, Unit 3043, University of Connecticut, Storrs, Connecticut 06269 USA
Received for publication 19 October 2007. Accepted for publication 7 April 2008.
ABSTRACT
Japanese barberry (Berberis thunbergii DC.) is a widespread invasive plant that remains an important landscape shrub represented by ornamental, purple-leaved forms of the botanical variety atropurpurea. These forms differ greatly in appearance from feral plants, bringing into question whether they contribute to invasive populations or whether the invasions represent self-sustaining populations derived from the initial introduction of the species in the late 19th century. In this study we used amplified fragment length polymorphism (AFLP) markers to determine whether genetic contributions from B. t. var. atropurpurea are found within naturalized Japanese barberry populations in southern New England. Bayesian clustering of AFLP genotypes and principal coordinate analysis distinguished B. t. var. atropurpurea genotypes from 85 plants representing five invasive populations. While a single feral plant resembled B. t. var. atropurpurea phenotypically and fell within the same genetic cluster, all other naturalized plants sampled were genetically distinct from the purple-leaved genotypes. Seven plants from two different sites possessed morphology consistent with Berberis vulgaris (common barberry) or B. xottawensis (B. thunbergii x B. vulgaris). Genetic analysis placed these plants in two clusters separate from B. thunbergii. Although the Bayesian analysis indicated some introgression of B. t. var. atropurpurea and B. vulgaris, these genotypes have had limited influence on extant feral populations of B. thunbergii.
Key Words: AFLP • Bayesian analysis • Berberidaceae • Berberis thunbergii • Berberis thunbergii var. atropurpurea • invasive plants • molecular markers
Since its introduction to New England as a horticultural plant in the late 1800s (Dirr, 1998![]() ), Japanese barberry (Berberis thunbergii DC.) has naturalized in more than 30 states across the United States (Silander and Klepeis, 1999
), Japanese barberry (Berberis thunbergii DC.) has naturalized in more than 30 states across the United States (Silander and Klepeis, 1999![]() ; USDA, NRCS, 2007
; USDA, NRCS, 2007![]() ). Feral plants are established primarily from seed, which is dispersed by birds and small rodents (Silander and Klepeis, 1999
). Feral plants are established primarily from seed, which is dispersed by birds and small rodents (Silander and Klepeis, 1999![]() ; Decker et al., 1991
; Decker et al., 1991![]() ). Invasive populations of Japanese barberry may displace native flora (Clark et al., 1998
). Invasive populations of Japanese barberry may displace native flora (Clark et al., 1998![]() ) and alter soil chemistry, microbial communities, and earthworm populations (Ehrenfeld et al., 2001
) and alter soil chemistry, microbial communities, and earthworm populations (Ehrenfeld et al., 2001![]() ). The sale of B. thunbergii is banned in Massachusetts (MDAR, 2005
). The sale of B. thunbergii is banned in Massachusetts (MDAR, 2005![]() ) and New Hampshire (New Hampshire General Court, 2004
) and New Hampshire (New Hampshire General Court, 2004![]() ) and many other states across the United States are considering similar prohibitions.
) and many other states across the United States are considering similar prohibitions.
Most Japanese barberry plants sold today have purple foliage and are the botanical variety atropurpurea. Purple barberries constitute a $5 million dollar retail crop in Connecticut and are worth many millions nationwide. Legal bans on the sale of B. thunbergii will result in economic consequences for the nursery industry because invasive plant legislation does not distinguish between a species and its botanical varieties. Horticultural genotypes of B. t. var. atropurpurea differ in appearance from feral plants (Dirr, 1998![]() ), bringing into question whether they contribute to invasive populations. Although purple barberries may produce seedlings that resemble naturalized plants (Lehrer et al., 2006a
), bringing into question whether they contribute to invasive populations. Although purple barberries may produce seedlings that resemble naturalized plants (Lehrer et al., 2006a![]() ), whether they make significant contributions to feral populations remains unclear. For this reason, genetic analyses that allow us to assess the level of genetic connection between B. t. var. atropurpurea and invasive populations are necessary to determine if purple-leaved forms of Japanese barberry should be included in bans of the species.
), whether they make significant contributions to feral populations remains unclear. For this reason, genetic analyses that allow us to assess the level of genetic connection between B. t. var. atropurpurea and invasive populations are necessary to determine if purple-leaved forms of Japanese barberry should be included in bans of the species.
Fingerprinting using molecular marker techniques such as AFLP, RAPD, and RFLP has proven especially valuable for inferring the origin and population structure of invasive plants (Mueller and Wolfenbarger, 1999![]() ). For example, Midori et al. (2006) were able to determine using RAPDs that more than 80% of the Hedera helix invasions in the Pacific Northwest were derived from the popular cultivar ‘Hibernica.’ AFLP markers have been successfully used to distinguish among cultivars in many other cultivated species, including members of the woody genera Ulmus (Pooler and Townsend, 2005
). For example, Midori et al. (2006) were able to determine using RAPDs that more than 80% of the Hedera helix invasions in the Pacific Northwest were derived from the popular cultivar ‘Hibernica.’ AFLP markers have been successfully used to distinguish among cultivars in many other cultivated species, including members of the woody genera Ulmus (Pooler and Townsend, 2005![]() ), Stewartia (Nair et al., 2005
), Stewartia (Nair et al., 2005![]() ), and Cornus (Smith et al., 2007
), and Cornus (Smith et al., 2007![]() ). Most importantly, the utility of AFLP markers to identify and differentiate specific genotypes of B. thunbergii has been well documented (Cote and Leduc, 2007
). Most importantly, the utility of AFLP markers to identify and differentiate specific genotypes of B. thunbergii has been well documented (Cote and Leduc, 2007![]() ;Lubell et al., 2008
;Lubell et al., 2008![]() ).
).
Our study explored whether a genetic link can be established between B. t. var. atropurpurea and invasive populations through the use of AFLP markers. Although AFLPs are dominant markers, complicating some aspects of the analysis, they are a widely adopted marker system in plants because they rapidly provide a large number of markers, are highly reproducible, and require no prior genetic information (Vos et al., 1995![]() ; Jones et al., 1997
; Jones et al., 1997![]() ).
).
MATERIALS AND METHODS
Plant material
In spring 2004 leaf tissue was collected for DNA extraction and AFLP analysis from 85 feral Japanese barberry plants within five New England invasive populations (Table 1), seven horticultural genotypes of B. t. var. atropurpurea, one cultivated genotype of B. vulgaris, and three cultivated B. xottawensis hybrids (Table 2). The invasive populations were located at Beaver Brook Park in Waltham, Massachusetts (42�22' N, –71�14' E); Bluff Point State Park in Groton, Connecticut (41�21' N, –72�04' E); Kent Falls State Park in Kent, Connecticut (41�43' N, –73�28' E); University of Connecticut Forest, Pink Ravine Preserve in Mansfield, Connecticut (41�48' N, –72�14' E); and Spring Manor Farm in Mansfield, Connecticut (41�48' N, –72�14' E) (Table 1). Cultivated plant material was derived from plants located at the University of Connecticut Plant Science Research Farm (Storrs, Connecticut) and in New England landscape situations (Table 2). Leaves collected in the field were held on ice before being stored at –80�C until DNA extraction.
|
|
Data analysis
Markers were scored initially using Genotyper 3.6 software (Applied Biosystems, Foster City, California, USA). Each marker identified by the software was visually confirmed before being assigned a value of 1 (presence of a marker) or 0 (absence of a marker) for each plant. A similarity matrix based on Jaccard’s coefficient of similarity was constructed using the SIMQUAL function in NTSYSpc 2.2 software (Exeter Software, Setauket, New York) (Rohlf, 2005![]() ). The DCENTER and EIGEN functions were used to perform principal coordinate (PCOORD) analysis and derive a two-dimensional ordination. Individuals were also clustered into conceptual populations based on their genotype using a Bayesian classification scheme as implemented in the software program Structure 2.0 (Pritchard et al., 2000
). The DCENTER and EIGEN functions were used to perform principal coordinate (PCOORD) analysis and derive a two-dimensional ordination. Individuals were also clustered into conceptual populations based on their genotype using a Bayesian classification scheme as implemented in the software program Structure 2.0 (Pritchard et al., 2000![]() ). The software was run for 10 iterations using a burnin period of 5000 and a sample of 25000 in the Markov chain Monte Carlo simulations. Our analyses considered numbers of conceptual populations ranging from two to 10. We determined that the optimal number of populations was three following analysis using the methods of Pritchard et al. (2000)
). The software was run for 10 iterations using a burnin period of 5000 and a sample of 25000 in the Markov chain Monte Carlo simulations. Our analyses considered numbers of conceptual populations ranging from two to 10. We determined that the optimal number of populations was three following analysis using the methods of Pritchard et al. (2000)![]() and Evanno et al. (2005
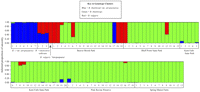
and Evanno et al. (2005![]() ; Table 3). The method of Pritchard et al. is based on the maximum natural logarithm of the probability of the data, and the method of Evanno et al. is based on the rate of change in the log probability of data between successive conceptual populations. Structure results are presented in a bar chart where each column corresponds to one individual plant.
; Table 3). The method of Pritchard et al. is based on the maximum natural logarithm of the probability of the data, and the method of Evanno et al. is based on the rate of change in the log probability of data between successive conceptual populations. Structure results are presented in a bar chart where each column corresponds to one individual plant.
|
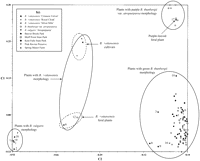
The six selective primer pairs produced 180 markers of which 147 were polymorphic. Structure analysis identified three genotype clusters, which are displayed in different colors in Fig. 1. The blue color indicates B. t. var. atropurpurea genotypes, the green color indicates green B. thunbergii genotypes, and the red color indicates B. vulgaris genotypes. Both Structure and PCOORD analysis assigned B. t. var. atropurpurea accessions to a unique cluster separate from green B. thunbergii plants (Figs. 1 and 2). One feral plant sampled from the Beaver Brook Park population (plant 17; Figs. 1 and 2) however, had purple foliage similar to B. t. var. atropurpurea genotypes and fell within the Structure and PCOORD analysis cluster for B. t. var. atropurpurea, suggesting that this plant may be a seedling derived from nearby cultivated specimens of B. t. var. atropurpurea. This result is not surprising because Beaver Brook Park lies within a heavily populated urban area and purple-leaved forms of Japanese barberry are abundant in the landscape.
|
|
Several additional feral plants were identified by Structure as having 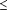 5% posterior probability of membership in the B. t. var. atropurpurea cluster (Fig. 1). PCOORD analysis suggests that none of these individuals have substantial B. t. var. atropurpurea influence in their ancestry. Detection of the purple-leaved feral plant at Beaver Brook Park and the introgression at Kent Falls State Park and Pink Ravine Preserve do reveal that B. t. var. atropurpurea genotypes can contribute their genes to invasive populations. Nevertheless, almost all other naturalized barberry plants sampled clustered with feral green-leaved B. thunbergii (Figs. 1 and 2), suggesting that the direct recruitment of seedlings from purple forms is a rare event.
5% posterior probability of membership in the B. t. var. atropurpurea cluster (Fig. 1). PCOORD analysis suggests that none of these individuals have substantial B. t. var. atropurpurea influence in their ancestry. Detection of the purple-leaved feral plant at Beaver Brook Park and the introgression at Kent Falls State Park and Pink Ravine Preserve do reveal that B. t. var. atropurpurea genotypes can contribute their genes to invasive populations. Nevertheless, almost all other naturalized barberry plants sampled clustered with feral green-leaved B. thunbergii (Figs. 1 and 2), suggesting that the direct recruitment of seedlings from purple forms is a rare event.
Following the introduction of Japanese barberry in Boston, leading horticulturists including Charles Springer Sargent (1889)![]() recognized its ornamental potential, and B. thunbergii quickly entered commercial production through local nurseries. As part of its 1895 offering, for example, Old Colony Nurseries (Plymouth, MA) advertised the availability of 10000 0.46–0.61 m tall Japanese barberry shrubs (Anonymous, 1895
recognized its ornamental potential, and B. thunbergii quickly entered commercial production through local nurseries. As part of its 1895 offering, for example, Old Colony Nurseries (Plymouth, MA) advertised the availability of 10000 0.46–0.61 m tall Japanese barberry shrubs (Anonymous, 1895![]() ). Herbarium records and botanist reports indicate that by the 1910s, B. thunbergii was found as a garden escape near Boston and adjacent to vacation homes maintained by Boston residents in the Berkshires, Nantucket, and elsewhere (Silander and Klepeis, 1999
). Herbarium records and botanist reports indicate that by the 1910s, B. thunbergii was found as a garden escape near Boston and adjacent to vacation homes maintained by Boston residents in the Berkshires, Nantucket, and elsewhere (Silander and Klepeis, 1999![]() ). From this initial foothold, Japanese barberry derived from the Boston introduction became firmly established in southern New England and by the 1920s "had spread in concentric circles" around the city (Silander and Klepeis, 1999
). From this initial foothold, Japanese barberry derived from the Boston introduction became firmly established in southern New England and by the 1920s "had spread in concentric circles" around the city (Silander and Klepeis, 1999![]() , p. 193). By the 1930s Japanese barberry was well established throughout much of southern New England and adjacent areas.
, p. 193). By the 1930s Japanese barberry was well established throughout much of southern New England and adjacent areas.
Feral green Japanese barberry populations were already well established by the time ornamental B. t. var. atropurpurea entered American cultivation in 192 6 (Dirr, 1998![]() ). Despite the presence of B. t. var. atropurpurea for
). Despite the presence of B. t. var. atropurpurea for  75 yr in southern New England, a period substantially greater than the time required for the original Boston introductions to become widespread, this purple-leaved form appears to have had minimal influence on feral populations.
75 yr in southern New England, a period substantially greater than the time required for the original Boston introductions to become widespread, this purple-leaved form appears to have had minimal influence on feral populations.
The inability of the widely grown purple-leaved Japanese barberry genotypes to substantially influence feral populations in recent decades may be attributed to several factors including reduced seed production by purple barberries and reforestation that affects seedling establishment. Lehrer et al. (2006b)![]() studied the reproductive potential of ornamental B. thunbergii genotypes and found that some purple barberries had reduced fruit and seed production relative to feral forms. This limited fecundity may impede the establishment of B. t. var. atropurpurea offspring in natural areas. Densely shaded woodlands, resulting from widespread reforestation in the second half of the 20th century may have also impeded the recruitment of progeny. The original spread of Japanese barberry throughout southern New England coincided with early reforestation after the abandonment of farms in the late 19th century (Spurr and Cline, 1942
studied the reproductive potential of ornamental B. thunbergii genotypes and found that some purple barberries had reduced fruit and seed production relative to feral forms. This limited fecundity may impede the establishment of B. t. var. atropurpurea offspring in natural areas. Densely shaded woodlands, resulting from widespread reforestation in the second half of the 20th century may have also impeded the recruitment of progeny. The original spread of Japanese barberry throughout southern New England coincided with early reforestation after the abandonment of farms in the late 19th century (Spurr and Cline, 1942![]() ; Silander and Klepeis, 1999
; Silander and Klepeis, 1999![]() ; Foster and O’Keefe, 2000
; Foster and O’Keefe, 2000![]() ). Such highly disturbed, lightly shaded understory conditions are optimal for barberry seedling establishment (Ehrenfeld et al., 2001
). Such highly disturbed, lightly shaded understory conditions are optimal for barberry seedling establishment (Ehrenfeld et al., 2001![]() ; Cassidy et al., 2004
; Cassidy et al., 2004![]() ). By the time purple barberries began to be commonly used in the 1930s and 1940s, forests had matured (Foster and O’Keefe, 2000
). By the time purple barberries began to be commonly used in the 1930s and 1940s, forests had matured (Foster and O’Keefe, 2000![]() ) and produced denser shade in the understory, reducing the survival of B. thunbergii seedlings (Silander and Klepeis, 1999
) and produced denser shade in the understory, reducing the survival of B. thunbergii seedlings (Silander and Klepeis, 1999![]() ).
).
Molecular analysis also detected the influence of B. vulgaris L. (common barberry) within four of the study populations (Fig. 1). Common barberry has been present in the New England environment since the colonial period (Steffey, 1985![]() ) and persists today as sporadic feral plants, despite an eradication campaign mounted in the first half of the 20th century (Peterson et al., 2005
) and persists today as sporadic feral plants, despite an eradication campaign mounted in the first half of the 20th century (Peterson et al., 2005![]() ). Visual inspection of four plants from the Beaver Brook Park population (plants 1, 2, 18 and 19; Fig. 1) and one plant from the Bluff Point State Park population (plant 1) revealed the larger leaves, serrated leaf margins, and racemose inflorescences typical of B. vulgaris (Krussmann, 1984
). Visual inspection of four plants from the Beaver Brook Park population (plants 1, 2, 18 and 19; Fig. 1) and one plant from the Bluff Point State Park population (plant 1) revealed the larger leaves, serrated leaf margins, and racemose inflorescences typical of B. vulgaris (Krussmann, 1984![]() ). Structure and PCOORD analysis confirmed this observation by aligning these plants with B. vulgaris ‘Atropurpurea’ in a cluster separate from the B. thunbergii group (Figs. 1 and 2).
). Structure and PCOORD analysis confirmed this observation by aligning these plants with B. vulgaris ‘Atropurpurea’ in a cluster separate from the B. thunbergii group (Figs. 1 and 2).
One plant from the Beaver Brook Park population (plant 6; Figs. 1 and 2) and one plant from the Bluff Point State Park population (plant 12) had entire leaf margins and subumbellate racemes characteristic of B. xottawensis (B. thunbergii x B. vulgaris) (Mehrhoff et al., 2007![]() ). PCOORD analysis supported this hybrid identity by placing these plants in a cluster between the groupings for B. vulgaris types and green-leaved B. thunbergii (Fig. 2). These two putative hybrids lie in the same plane as three ornamental barberry cultivars (‘Crimson Velvet,’ ‘Royal Cloak,’ and ‘Silver Mile’) that possess similar morphology and are reported to be selections of B. xottawensis (Houtman, 2004
). PCOORD analysis supported this hybrid identity by placing these plants in a cluster between the groupings for B. vulgaris types and green-leaved B. thunbergii (Fig. 2). These two putative hybrids lie in the same plane as three ornamental barberry cultivars (‘Crimson Velvet,’ ‘Royal Cloak,’ and ‘Silver Mile’) that possess similar morphology and are reported to be selections of B. xottawensis (Houtman, 2004![]() ; Lubell et al., 2008
; Lubell et al., 2008![]() ; Fig. 2). Within this cluster, the three cultivars fall closer to the grouping for purple-leaved B. t. var. atropurpurea, while the two feral seedlings are situated closer to the grouping for green-leaved B. thunbergii. These positions are significant because the B. xottawensis cultivars possess purple foliage likely derived from a B. t. var. atropurpurea parent, whereas the feral B. x ottawensis seedlings are green. Structure analysis supports the hybrid parentage of ‘Crimson Velvet,’ ‘Royal Cloak,’ and ‘Silver Mile’ by showing equal contributions for B. t. var. atropurpurea and B. vulgaris (Fig. 1). Similarly, structure analysis shows feral B. xottawensis plants as half green B. thunbergii and half B. vulgaris (Fig. 1).
; Fig. 2). Within this cluster, the three cultivars fall closer to the grouping for purple-leaved B. t. var. atropurpurea, while the two feral seedlings are situated closer to the grouping for green-leaved B. thunbergii. These positions are significant because the B. xottawensis cultivars possess purple foliage likely derived from a B. t. var. atropurpurea parent, whereas the feral B. x ottawensis seedlings are green. Structure analysis supports the hybrid parentage of ‘Crimson Velvet,’ ‘Royal Cloak,’ and ‘Silver Mile’ by showing equal contributions for B. t. var. atropurpurea and B. vulgaris (Fig. 1). Similarly, structure analysis shows feral B. xottawensis plants as half green B. thunbergii and half B. vulgaris (Fig. 1).
The Structure analysis provided additional evidence of B. vulgaris introgression at the Beaver Brook Park, Kent Falls State Park, and Pink Ravine Preserve populations. One feral plant at Beaver Brook Park (plant 3; Fig. 1) had a 13% posterior probability of membership within the B. vulgaris cluster. Similarly, one feral plant at Pink Ravine Preserve (plant 16) had a 6%, and two plants at Kent Falls State Park (plants 8 and 9) had 18% and 14% posterior probabilities of membership within the B. vulgaris cluster. The PCOORD analysis places plants 3, 8 and 9 closer to the feral B. xottawensis cluster than it does the other green-leaved feral B. thunbergii plants. Plant 16 did not separate from the green-leaved B. thunbergii genotypes (Fig. 2).
While the majority of feral plants sampled across all five populations had genotypes and morphology consistent with B. thunbergii, the results of our study indicate that common barberry and purple barberry have exerted some genetic influence on extant invasive barberry populations in southern New England. Our findings provide some insight into how two invasive species (B. thunbergii and B. vulgaris) can interact in invasive populations and also how a horticultural variety can behave as a component of an invasive landscape. While our results suggest that the contribution of horticultural varieties to feral populations is small, we detect evidence of that influence even in some individuals that lack the obvious visual traits associated with purple-leaved forms. Thus, our results suggest that local, state, and national authorities should consider the risk that sale of purple-leaved Japanese barberries will inadvertently contribute to existing feral populations.
FOOTNOTES
4 Author for correspondence (e-mail: Mark.Brand@uconn.edu) ![]()
5 Present address: Department of Ornamental Horticulture, Farmingdale State College, Farmingdale, New York 11735 USA ![]()
LITERATURE CITED
Anonymous. 1895. Advertisement. Garden and Forest 8: 2.
Cassidy, T. M., J. H. Fownes, AND R. A. Harrington. 2004. Nitrogen limits an invasive perennial shrub in forest understory. Biological Invasions 6: 113–121.[CrossRef][ISI]
Cheng, F. S., S. K. Brown, AND N. F. Nelson. 1997. A DNA extraction protocol from various tissues in woody species. HortScience 32: 921–922.
Clark, F. H., C. Mattrick, AND S. Shonbrun. 1998. Rogues gallery: New England’s notable invasives. Conservation Notes of the New England Wildflower Society 2: 22.
Clarke, M. M., S. H. Reichard, AND C. W. Hamilton. 2006. Prevalence of different horticultural taxa of ivy (Hedera spp., Araliaceae) in invading populations. Biological Invasions 8: 149–157.[CrossRef][ISI]
Cote, M., AND L. Leduc. 2007. Molecular identification of Japanese barberry (Berberis thunbergii) cultivars using amplified fragment length polymorphism. HortScience 42: 478–482.
Decker, S. R., P. J. Pekins, AND W. M. Mautz. 1991. Nutritional evaluation of winter foods of wild turkeys. Canadian Journal of Zoology 69: 2128–2132.
Dirr, M. A. 1998. Manual of woody landscape plants. Stipes Publishing, Champaign, Illinois, USA.
Ehrenfeld, J. G., P. Kourtev, AND W. Huang. 2001. Changes in soil functions following invasions of exotic understory plants in deciduous forests. Ecological Applications 11: 1287–1300.[CrossRef][ISI]
Evanno, G., S. Regnaut, AND J. Goudet. 2005. Detecting the number of clusters of individuals using the software structure: A simulation study. Molecular Ecology 14: 2611–2620.[CrossRef][Medline]
Foster, D. R., AND J. F. O’Keefe. 2000. New England forests through time. Harvard University Press, Cambridge, Massachusetts, USA.
Houtman, R. 2004. Variegated trees and shrubs. Timber Press, Portland, Oregon, USA.
Jones, C. J., K. J. Edwards, S. Castaglione, M. O. Winfield, F. Sala, C. van de Wiel, G. Bredemeijer, B. Vosman, M. Matthes, A. Daly, R. Brettschneider, P. Bettini, M. Buiatte, E. Maestri, A. Malcevschi, N. Marmiroli, R. Aert, G. Volckaert, J. Rueda, R. Linacero, A. Vazques, AND A. Karp. 1997. Reproducibility testing of RAPD, AFLP and SSR markers in plants by a network of European laboratories. Molecular Breeding 3: 381–390.[CrossRef][ISI]
Krussmann, G. 1984. Manual of cultivated broad-leaved trees and shrubs, vol. 1, A–D. Timber Press, Portland, Oregon, USA.
Lehrer, J. M., M. H. Brand, AND J. D. Lubell. 2006a. Seedling populations produced by colored-leaf genotypes of Japanese barberry (Berberis thunbergii DC.) contain seedlings with green leaf phenotype. Journal of Environmental Horticulture 24: 133–136.
Lehrer, J. M., M. H. Brand, AND
J. D. Lubell. 2006b. Four cultivars of Japanese barberry demonstrate
differential reproductive potential under landscape conditions. HortScience 41: 762–767.
Lubell, J. D., M. H. Brand, AND J. M. Lehrer. 2008. AFLP identification of Berberis thunbergii cultivars, inter-specific hybrids, and their parental species. Journal of Horticultural Science &Biotechnology 83: 55–63.[ISI]
MDAR [Massachusetts Department of Agricultural Resources]. 2005. Massachusetts prohibited plant list. MDAR, Boston, Massachusetts, USA. Website http://mass.gov/agr/farmproducts/Prohibited_Plant_Index2.htm [accessed November 2007].
Mehrhoff, L. J., J. A. Silander Jr., S. A. Leicht, E. S. Mosher, AND N. M. Tabak. 2007. IPANE: Invasive plant atlas of New England. Website http://nbii-nin.ciesin.columbia.edu/ipane [accessed 25 July 2007].
Mueller, U. G., AND L. Wolfenbarger. 1999. AFLP genotyping and fingerprinting. Trends in Ecology &Evolution 14: 389–394.[CrossRef][ISI][Medline]
Nair, A., D. Zhang, AND D. Hu. 2005. Application of AFLP markers on taxon discrimination of cultivated Stewartia. HortScience 40: 1041.
New Hampshire General Court. 2004. Adopted Rule, Chapter Agr 3800, Invasive species. In State of New Hampshire, New Hampshire code of administrative rules. Website http://www.nh.gov/agric/divisions/plant_industry/documents/Rules_8.pdf [Accessed November 2007].
Perry, M. D., M. R. Davey, J. B. Power, K. C. Lowe, H. F. J. Bligh, P. S. Roach, AND C. Jones. 1998. DNA isolation and AFLP genetic fingerprinting of Theobroma cacao (L.) [sic]. Plant Molecular Biology Reporter 16: 49–59.[CrossRef][ISI]
Peterson, P. D., K. J. Leonard, J. D. Miller, R. J. Laudon, AND T. B. Sutton. 2005. Prevalence and distribution of common barberry, the alternate host of Puccinia graminis, in Minnesota. Plant Disease 89: 159–163.[CrossRef][ISI]
Pooler, M. R., AND A. M. Townsend. 2005. DNA fingerprinting of clones and hybrids of American elm and other elm species with AFLP markers. Journal of Environmental Horticulture 23: 113–117.
Pritchard, J. K., M. Stephens, AND P. Donnelly. 2000. Inference of population structure using multilocus genotype data. Genetics 155: 945–959.
Rohlf, J. F. 2005. NTSYSpc. Applied Biostatistics, Port Jefferson, New York, USA.
Sargent, C. C. 1889. New or little known plants. Garden and Forest 2: 52.
Silander, J. A., AND D. M. Klepeis. 1999. The invasion ecology of Japanese barberry (Berberis thunbergii) in the New England landscape. Biological Invasions 1: 189–201.[CrossRef]
Smith, N. R., R. N. Trigiano, M. T. Windham, K. H. Lamour, L. S. Finley, X. Wang, AND T. A. Rinehart. 2007. AFLP markers identify Cornus florida cultivars and lines. Journal of the American Society for Horticultural Science 132: 90–96.
Spurr, S. H., AND A. C. Cline. 1942. Ecological forestry in central New England. Journal of Forestry 40: 418–420.
Steffey, J. 1985. Strange relatives: The barberry family. American Horticulturalist 64: 4–9.
USDA, NRCS [U.S. Department of Agriculture, Natural REsources Conservation Service]. 2007. The PLANTS database, version 3.5, National Plant Data Center, Baton Rouge, Louisiana, USA. Website http://plants.usda.gov [Accessed 5 May 2007].
Vos, P., R. Hogers, M. Bleeker, M. Reijans, T. van de Lee, M. Hornes, A. Fijters, J. Pot, J. Peleman, M. Kuiper, AND M. Zabeau. 1995. AFLP: A new technique for DNA fingerprinting. Nucleic Acids Research 23: 4407–4414.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| HOME | HELP | FEEDBACK | SUBSCRIPTIONS | ARCHIVE | SEARCH | TABLE OF CONTENTS |