Batrachotoxin Holly Grimes |
 Phyllobates
terribilis:
a batrachotoxin-containing poison dart frog. Phyllobates
terribilis:
a batrachotoxin-containing poison dart frog.(photograph from Jensen (12)) |
Abstract
Batrachotoxin (BTX), a member of the batrachotoxin family of steroidal alkaloids, was first isolated from the skin of poison arrow frogs (genus Phyllobates) from the rain forests of western Colombia. This neurotoxin has also been found in Choresine beetles; in addition, BTX has been synthesized in the laboratory from (+/-)-cis-decalone. In animals, batrachotoxin inactivates the sodium channels in nerve and muscle cells, thereby interfering with the electrical signals sent throughout the body and causing fibrillation, arrhythmias, cardiac failure, and death. This potent toxin is used as a poison for blow darts and as an aid for research regarding voltage-gated sodium channels. In the future, batrachotoxin could be used as an active ingredient in pain killer ointments.
General Information
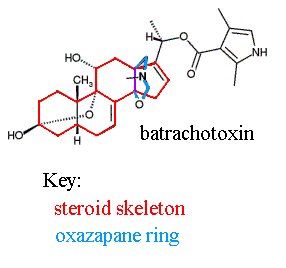
The neurotoxin batrachotoxin (BTX) is a member of a family of steroidal alkaloids called batrachotoxins. The batrachotoxin family includes (among other chemicals) batrachotoxin, homobatrachotoxin, batrachotoxinin A, and pseudobatrachotoxin (unstable, converts to batrachotoxinin A on standing) (7). These toxins were first discovered in poison dart frogs of the genus Phyllobates (8); the name given to this group of chemicals was derived from the greek word for frog, "batrachos" (9). Batrachotoxin, the (20-alpha)-2,4-dimethylpyrrole-3-carboxylate of batrachotoxinin-A, can be assayed using a modified Ehrlich reagent (detection limit less than 50 ng) (7); Ehrlich reagent is an acid solution of p-dimethyl amino benzaldehyde (10). The structure of BTX includes a steroid skeleton and an oxazapane ring (13), which are indicated on the structure below.
By weight, batrachotoxin is one of the most potent natural toxins known (9) because it binds to and irreversibly opens voltage-gated sodium channels (9,15); the proper function of these sodium channels is required for the transmission of electrical signals through nerve and muscle cells (2). The activity of BTX depends on temperature, reaching its maximum at 37 degrees Celsius (4). The following list gives some of the properties of this potent toxin.
Other Names (3): BTX; Batrachotoxin (7CI, 8CI); Batrachotoxinin A, 20-(2,4-dimethyl-1H-pyrrole-3-carboxylate); 1H-Pyrrole-3-carboxylic acid, 2,4-dimethyl-, (1S)-1-[(5aR, 7aR, 9R, 11aS, 11bS, 12R, 13aR)-1,2,3,4,7a,8,9,10,11,11a,12,13-dodecahydro-9,12-dihydroxy-2,11a-dimethyl-7H-9,11b-epoxy-13a,5a-propenophenanthro[2,1-f] [1,4] oxazepin-14-yl] ethyl ester (9CI) (CA index name); 1H-Pyrrole-3-carboxylic acid, 2,4-dimethyl-, 1-(1,2,3,4,7a,8,9,10,11,11a,12,13-dodecahydro-9,12-dihydroxy-2,11a-dimethyl-7H-9,11b-epoxy-13a,5a-propenophenanthro[2, 1-f] [1,4] oxazepin-14-yl) ethylester, [5aR- [5aa, 7ab, 9a, 11ab, 11ba, 12a, 13aa, 14 (S*)]]-
| Molecular Formula
(3): C31 H42 N2 O6 Registry Number (3): 23509-16-2 Predicted Boiling Point (3): 744.0 +/- 60.0 deg. C Predicted Density (3): 1.34 +/- 0.1 g/cm^3 Predicted pKa (25 deg C) (3): 7.87 +/- 0.70 (most basic) to 12.93 +/- 0.70 (most acidic) LD50 in mice (1): 2 µg/kg |
|
Discovery
In the 1960s, John Daly and his collegues collected samples of toxin from the skin of Phyllobates aurotaenia (a poison-dart frog)and isolated from those samples four poispns that they would later name batrachotoxin, homobatrachotoxin, batrachotoxinin A, and pseudobatrachotoxin (7). Unfortunately, only small quantities of batrachotoxin (50 µg (1)) could be collected from each P. aurotaenia specimen, limiting the amount of toxin available for use in research. This supply problem was solved when it was discovered that P. terribilis provided a much greater quantity of BTX per individual (1000 µg per frog (8)) than did any other frog known to possess this toxin (14).
The batrachotoxins were first isolated from poison samples by the following procedure. Methanolic extracts from the P. aurotaenia frog skins were first concentrated at low pressure, and then partitioned between chloroform and water. Next, the basic components of the skin extracts (present in the chloroform layer) were extracted into 0.1 N HCl. These extracts were basified using 1N NH4OH and then reextracted into chloroform. The batrachotoxins were then purified using preparative thin-layer or column chromatography using silica gel (1).
Natural Sources
Batrachotoxin (along with homobatrachotoxin, batrachotoxinin-A, and pseudobatrachotoxin) is secreted from glands in the skin of poison-dart frogs of the genus Phyllobates from the rain forests of western Colombia (4, 8). Of the frogs that carry the toxin, P. terribilis, P. bicolor, and P. aurotaenia possess large amounts of batrachotoxin (8). These frogs do not produce the toxins themselves: when raised in captivity with no alkaloids in their diets, Phyllobate poison-dart frogs do not possess detectable concentrations of batrachotoxins (8, 9). Thus, P. terribilis and its relatives probably obtain the toxins from their diet--possibly from Coresine beetles, which are known to contain batrachotoxins in high concentrations (Coresine beetles most likely also provide the homobatrachotoxin found in the passerine birds of New Guinea). Current evidence indicates that beetles cannot synthesize steroid skeletons (such as the one in batrachotoxin) de novo, so the insects probably obtain these skeletons from the phytosterols (such as cholesterol) in the plants they eat (9). Poison-dart frogs are able to sequester these toxins without being killed by them because the frogs have evolved voltage-gated sodium channels that are unaffected by the toxin (7).
Synthesis
|
Before 1998, it was thought that batrachotoxins were so complex that they could not be synthesized in the laboratory (14). Batrachotoxin, at that time, had only been partially synthesized from batrachotoxinin A (1). However, in 1998, Kurosu et al reported that they had synthesized (+/-)-Batrachotoxinin A from (+/-)-cis-decalone; this synthesis can be considered a "formal total synthesis" of (+/-)-batrachotoxin since batrachotoxinin A can be converted into batrachotoxin (13). Important steps in this synthesis include an intramolecular furan Diels-Alder reaction to construct the steroid skeleton, and an intramolecular oxy-Michael reaction to synthesize the oxazapane ring (13). In general, the Diels-Alder reaction is used to form six-membered rings such as those present in steroid skeletons (17) and the Michael reaction uses carbonyl compounds as reactants (18) (these carbonyl compounds are intermediates and reagents involved in the synthesis of batrachotoxin (13)). |
|

Biological Effects
|
Contact with batrachotoxin causes numbness in human tissue. Upon entering the body, this toxin causes muscle and nerve depolarization, fibrillation, arrhythmias, and heart failure (5). BTX causes these deadly effects by interfering with the body's ability to transmit electrical signals by means of action potentials. Action potentials. All cells in the body have an electric potential difference of 70 to 80 mV across their outer membranes, with the inside of the cell being negatively charged with respect to the outside environment. This membrane potential is caused by the unequal distribution of ions across the membrane: sodium ions are more concentrated outside the cell while potassium ions are more concentrated inside. The movement of these ions across the cell membrane is controlled by voltage-gated sodium and potassium channels; when the cell is at rest, all of the sodium channels are closed while some of the potassium channels are open (2). From this resting state, action potentials are triggered by local depolarizations during which the inside of the cell becomes positive with respect to the outside; these depolarizations are initiated in nerve cells by the sense organs. When the depolarization exceeds the threshold voltage (usually 10 mV, but at most 50 mV), an action potential is initiated and the voltage-gated sodium channels open. The resulting inward flow of sodium ions depolarizes the cell even further (usually to a charge greater than 0.1 V). The cell subsequently repolarizes when the sodium channels enter their inactivated state (a special type of closed state) and the potassium channels open. Once repolarization is complete, the cell returns to the resting state. Action potentials last for less than 1 millisecond and involve the movement of a small number of ions; in the face of this ion movement, the sodium and potassium ion concentration gradients are maintained by a sodium-potassium pump (2). Batrachotoxin's influence. Batrachotoxin inhibits the ability of nerve and muscle cells to transmit these electrical signals by binding to voltage-gated sodium channels. In binding, BTX modifies the sodium channel so that the threshold voltage (the voltage at which the sodium channels open) shifts to more negative voltages, the channels cannot be inactivated, and the channels' ion selectivity is reduced (5, 15, 16). Tetrodotoxin (puffer fish poison (4)), local anesthetics, and certain anticonvulsants and antiarrhythmic drugs inhibit the binding of batrachotoxin (16). BTX can only bind to a sodium channel when the channel is in its open conformation, so it has been hypothesized that batrachotoxin binds within the pore of the channel (15). These channels are glycoproteins (11) and the ability of batrachotoxin to bind depends on the amino acid sequences of these proteins: BTX is stabilized within its receptor by the electrostatic interaction of batrachotoxin's positively charged (protonated) tertiary amine with the phenylalanine residue's aromatic group. Amino acid substitutions that prevent this type of electrostatic interaction inhibit the ability of BTX to bind to the sodium channels. Once bound, batrachotoxin shifts the equilibrium among the possible channel conformations (open, closed, and inactivated) to strongly favor the open state; this activation of the sodium channel is irreversible (15).
|
 |
 |
|
 |
|
 |


Uses
Batrachotoxin is used by the Embera Indians of western Colombia to poison their blow darts. Poison dart frogs of the genus Phyllobates (P. terribilis, P. bicolor, and P. aurotaenia) are the Indians' source for this toxin (7); simply scraping the back of one of these frogs with a dart provides sufficient poison to kill a monkey or a bird (14). BTX is also used extensively in research regarding voltage-gated sodium channels and these channels' structure, function, and interaction with ligands (6, 11, 15, 16). Lastly, batrachotoxin may one day be used as an active ingredient in pain killer ointments (5).
Unfortunately, to obtain toxin for the uses just listed, Phyllobate frogs must be injured or killed: the Indians hold the frogs over a fire or run sticks through the frogs' bodies so that the amphibians will secrete poison (4), and scientists must kill the frogs before extracting toxin from the skins (14).
Conclusions
Batrachotoxin has proven useful to humans as a blow dart poison, as a tool for sodium channel research, and (possibly) as an active ingredient in pain killers. Since BTX has been formally synthesized from a relatively simple starting material (cis-decalone), people may one day no longer need to depend on frogs of the genus Phyllobates as a major source of this toxin. Hopefully scientists will one day develop techniques for mass producing this interesting and useful toxin that do not involve killing or otherwise harming poison arrow frogs.
References
(1) Albuquerque, E.X.; Daly, J.W.; Witkop, B. Batrachotoxin: Chemistry and Pharmacology. Science 1971, 172, 995-1002.
(2) Ashcroft, F. Action Potentials. In The Oxford Companion to the Body; Blakemore, C.; Jennett, S., Eds.; In Oxford Reference Online; Oxford University Press, 2001; Western Oregon University. http://www.oxfordreference.com/views/ENTRY.html?subview=Main&entry=t128.e8 (1 March 2006).
(3) Batrachotoxin. RN 23509-16-2. STN Registry file (27 February 2006).
(4) Batrachotoxin. Wikipedia. http://en.wikipedia.org/wiki/Batrachotoxin (March 16 2006).
(5) Bosmans, F.; Maertens, C.; Verdonck, F.; Tytgat, J. The poison Dart frog's batrachotoxin modulates Nav1.8. FEBS Lett. 2004, 577, 245-248.
(6) Cronin, N.B.; O'Reilly, A.; Duclohier, H.; Wallace, B.A. Effects of Deglycosylation of Sodium Channels on Their Structure and Function. Biochemistry 2005, 44, 441-449.
(7) Daly, J.W. Thirty Years of Discovering Arthropod Alkaloids in Amphibian Skin. J. Nat. Prod. 1998, 61, 162-172.
(8) Daly, J.W.; Spande, T.F.; Garraffo, H.M. Alkaloids from Amphibian Skin: A Tabulation of Over Eight-Hundred Compounds. J. Nat. Prod. 2005, 68, 1556-1575.
(9) Dumbacher, J.P.; Wako, A.; Derrickson, S.R.; Samuelson, A.; Spande, T.F.; Daly, J.W. Melyrid beetles (Choresine): A putative source for the batrachotoxin alkaloids found in poison-dart frogs and toxic passerine birds. PNAS 2004, 101, 15857-15860.
(10) Ehrlich's Reagent. General Practice Notebook. http://www.gpnotebook.co.uk/cache/-872021970.htm (March 18 2006).
(11) Grauert, M.; Bechtel, W.D.; Weiser, T.; Stransky, W.; Nar, H.; Carter, A.J. Synthesis and Structure - Activity Relationships of 6,7-Benzomorphan Derivatives as Use-Dependent Sodium Channel Blockers for the Treatment of Stroke. J. Med. Chem. 2002, 45, 3755-3764.
(12) Jensen, J. Phyllobates Terribilis. Johnny Jensen's Photographic Library. http://www.jiphoto.dk/animal_archive/phyllobates_terribilis.htm (March 16 2006).
(13) Kurosu, M.; Marcin, L.R.; Grinsteiner, T.J.; Kishi, T. Total Synthesis of (+/-)-Batrachotoxinin A. J. Am. Chem. Soc. 1998, 120, 6627-6628.
(14) Lasley, E.N. Having Their Toxins and Eating Them Too. BioScience 1999, 49, 945-950.
(15) Li, H.; Hadid, D.; Ragsdale, D.S. The Batrachotoxin Receptor on the Voltage-Gated Sodium Channel is Guarded by the Channel Activation Gate. Mol. Pharmacol. 2002, 61, 905-912.
(16) Linford, N.J.; Cantrell, A.R.; Qy, Y.; Scheuer, T.; Catterall, W.A. Interaction of batrachotoxin with the local anesthetic receptor site in transmembrane segment IVS6 of the voltage-gater sodium channel. Proc. Natl. Acad. Sci. USA 1998, 95, 13947-13952.
(17) Smith, J.G. Organic Chemistry, 1st ed.; McGraw-Hill: New York, 2006; p 561-568.
(18) Smith, J.G. Organic Chemistry, 1st ed.; McGraw-Hill: New York, 2006; p 881.
(19) Smith, J.G. Organic Chemistry, 1st ed.; McGraw-Hill: New York, 2006; p 1014.